Luca Bondioli
University of Padua
Polish Academy of Sciences
When bioarchaeologists talk about age, they are referring to two things: chronological age and the biological markers that correspond to it. If I were one of the human skeletons that cross my lab bench, I would class myself into an age category which bioarchaeologists call ‘senile’. That is to say one which is well beyond the 50th year of life and within the last of the age classes that one assigns to a skeletal specimen (Buikstra and Ubelaker, 1994). The ‘senile’ age class clearly and unambiguously signifies that beyond the 50th year, there is no bio-cultural decay and degeneration that can be placed on the spectrum of normality. Nevertheless, I am more than my body, and the physical changes which my (well) over 50 years have earned me have also allowed me to accrue 40 years’ of experience as well. As a result of this, the organizers of the 30th EAA Annual Meeting in Rome asked me for a short a keynote speech, and this contribution is intended as a short summary of the perspectives that I presented on the occasion. This text represents some of the lessons I have learned from the discipline which I have been attempting to study, practice, teach, and even influence a little for more than forty years.
While I would argue that it is not for me to decide whether I have been successful in this decades-long endeavour of mine, I nevertheless firmly believe that the study of the bio-cultural aspects of our species (and our ancestors—both direct and cousins and relatives) is one of the most enjoyable undertakings one can employ in one's lifetime. Certainly, everything I am about to say is heavily influenced by my own cultural milieu: essentially that of an individual from a Western country, certainly weighed down by a very heavy colonial burden and distorted by the worst disciplinary histories imaginable (Mulligan, 2021, Schroeder, 2020).
Having thus made a minimal excusatio (non petita) as to why I am writing this text on biological anthropology, it remains to clarify at least in part how I came to think what I am about to illustrate.
Scientifically speaking, I was born in the 1970s into a world of anthropology. In particular, into the world of physical anthropology (as it was called then), where university classes were taught in the “biology of human races”. Such classes were not sporadic but were present in all Italian universities. Among the various disciplinary subfields, there was an attempt to catalogue humans by means of measurements and aspects of skull shape (I remember a conference of Italian anthropologists where—even then dismayed—I heard a speaker claim that the presence of a dolichocephalic skull among meso-brachycephals in a Sardinian tomb was “certain indication of migration and arrival of allochthons”). Thankfully, this world has disappeared and been somewhat forgotten. However, as I came from it, I nevertheless kept some of its best memories alive. These include a very strong fondness for the study of the human remains of our ancestors. With this in my heart, I had little inclination to leave the field behind. But fate, ever mocking, ultimately led me instead to think about data and statistics. I came to study the unseen, the microscopic, the compositional: to train my sights on isotopes and elemental concentrations. My work led me to focus on things that were not so tangible, but that could nevertheless produce a great deal of knowledge.
Since the 1990s, incredible technological advances have opened new windows into the past, including the study of ancient DNA, which in my scientific youth was an unattainable holy grail. With these leaps forward in mind, a necessary premise must first be made to put what follows into the correct perspective; we must consider the so-called ‘Osteological Paradox’.
The curse of using mortality samples and the ‘Osteological Paradox’
The study of past populations through the lens of bioarchaeology has always been divided between population approaches (i.e. analysing sufficiently numerous samples from funerary or catastrophic contexts and extracting parameters that can be traced back to the reference population) and individual approaches (studying skeletons, of parts of a skeleton, as separate, individual entities and trying to deduce bio-cultural aspects of the group to which they belonged). The former approach has certainly been more successful in that it has allowed (even in the paucity of analytical tools available until a few decades ago) the creation of effective bioarchaeological reconstructions. The latter approach, by contrast, has been confined to contexts with very few remains (typically the world of fossil hominins, with but a few exceptions) or to that part of palaeopathology that has seen articles of “a case of ...”. Unfortunately, the latter have often (but not always) had moderate impact on the advancement of knowledge. In this last, self-criticism is also needed as ‘qui sine peccato est vestrum, primus lapidem mittat’ (lit. ‘let he among you who is without sin cast the first stone’, John 8:2). I too am guilty in this respect, although on a very interesting case (Weaver et al., 2000).
This said, the great limitation of population-based bioarchaeological studies is that they are based on:
Mortality samples: barring the rare examples of assemblages from catastrophic events (Martyn et al., 2018, Martyn et al., 2020, Soncin et al., 2021), bioarchaeological remains come from funerary contexts thus represent a mirror (often distorted, see next point) of those who have died within a society, rather than reflecting who was alive. This simple consideration has never been sufficiently taken into account and has caused unrealistic interpretations. Indeed, mortality samples are deeply influenced by the so-called 'Osteological Paradox' (DeWitte and Stojanowski, 2015, Wood et al., 1992).
Wood et al. (1992) pointed out the three key issues that hinder our understanding of past health (more generally of bio-cultural adaptations): demographic non-stationarity, selective mortality and hidden heterogeneity in morbidity and mortality.
Demographic non-stationarity. Among demographic variables, migration has the effect of altering the profile of a population, since the health status and life history of newcomers do not correspond to those of the host community but are the product of different bio-cultural environments.
Hidden heterogeneity in the risk of disease and mortality. There are a multitude of factors that influence the probability of falling ill and dying. These run the gamut from genetic predisposition to acquired susceptibility. These factors and their interaction are unquantifiable, except in rare cases.
Selective mortality. In dealing with past populations, we should always remember that we are dealing with people who did not survive and thus likely embody higher frequencies of pathological conditions and other skeletal signs related to harsher lifestyles, such as growth disorders, poor nutritional intake, heavy workloads, etc. In some cases, human skeletal assemblages derived from catastrophic events can give us a glimpse into the living community, although they certainly cannot represent an exact cross-section of the population.
Non-random samples: in addition to what has been noted in point 1, it should be added that samples from funerary contexts are practically always non-random and are also subject to selection due to natural causes (e.g. only part of the necropolis preserved) and/or cultural causes (differential access to the necropolis for diverse segments of the living population, spatial segregation by groups, etc.) (Sperduti et al., 2021, Sperduti et al., 2018), thus further complicating the interpretative framework.
The ancient DNA revolution
A decade ago, Kristiansen (Kristiansen, 2014) rightly realised that the possibility we have today of reconstructing extended fragments of our ancestors' DNA and genetic profiles has set the stage for a true revolution in archaeology. Indeed, from a world of essentially morphological studies of the osteo-dental apparatus (human and animal), we are able today (with some limitations) to largely reconstruct the genetic heritage of our ancestors with considerable accuracy.
Initially, only the extraction and sequencing of non-sapiens DNA could be tackled (Prüfer et al., 2014, Prüfer et al., 2017, Green et al., 2010). But thanks to technological advances that will not be addressed here, it has been possible to extract and sequence endogenous DNA from bones and teeth from funerary and paleoanthropological contexts that also encompassed Homo sapiens (Reich, 2018). Today, the problem of modern DNA contamination that had so troubled the first decades of ancient DNA analyses has been contained (or at least controlled). A good summary of the current state of ancient DNA analyses can be found in Orlando et al. (2021). In their paper, they highlight how the process of extraction, sequencing, analysis and interpretation has reached such levels of technological and, particularly, bioinformatic complexity (specifically in relation to how extracting information and—even worse— independently elaborating the data) is almost incomprehensible to those outside of a genetics background.
That being said, geneticists and other researchers who produce data on the ancient (and modern) human genome follow strict rules of data sharing and promote the public availability of their results. All researchers deposit the raw DNA sequencing data in public repositories, such as ENA (Yuan et al., 2024), SRA (Kodama et al., 2012, Leinonen et al., 2011), GenBank (Benson et al., 2013), and others. Thus, any informed bioarchaeologist should be able to retrieve all the aDNA sequences for their sites of interest and analyze them. But there is still a deep gap between making data available and being able to work effectively with the enormous wealth of aDNA sequences out there. As a promising start in this sense, the Allen Ancient DNA Resource [AADR] (Mallick et al., 2024) crossed the threshold of >10,000 individuals in 2022 and may represent a hitherto unthinkable advance in the understanding of the adaptive dynamics of sapiens populations in the Holocene (Akbari et al., 2024). However, a user-friendly computer tool that allows anyone with a good background in palaeogenetics to use and analyse aDNA sequences has yet to materialise.
Lastly, the possibility of reconstructing recent genealogical connections between individuals intra and inter funerary contexts using long DNA segments shared between two individuals (known as identity-by-descent or IBD; Ringbauer et al., 2024), represents an incredible new exploratory tool adding a new metric to the problem of disentangling the mutual relationships between ancient communities. For more details, see Figure 1 (Ringbauer et al. 2024).
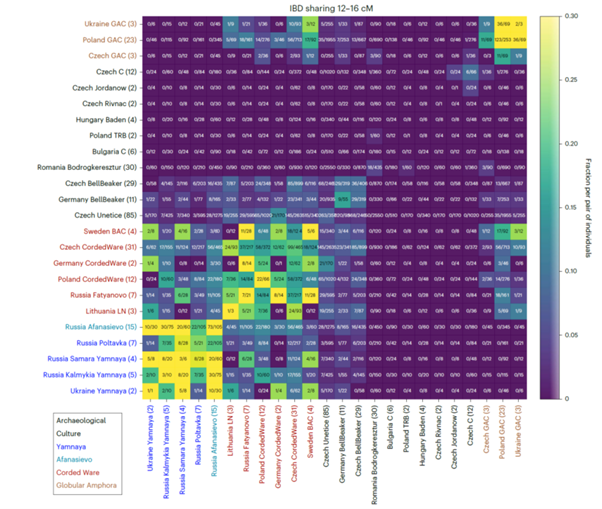
Figure 1. Original figure and caption from Ringbauer et al. (2024), figure 4: Inferred IBD segments between various Eneolithic and Bronze Age West Eurasian Groups. We visualize IBD segments 12–16 cM long. We applied ancIBD to identify IBD segments between all pairs of 304 West Eurasian ancient individuals (all previously published data) organized into 24 archaeological groups. The number in the parenthesis indicates the sample size for each archaeological group. For each pair of groups, we plot the fraction of all possible pairs of individuals that share at least one IBD 12–16 cM long, which we obtained by dividing the total number of pairs that share such IBD segments by the total number of all possible pairs: between two different groups of n1 and n2 individuals, one has n1n2 pairs, while within a group (on the diagonal in the figure) of size n one has n(n − 1)/2 pairs. LN, Late Neolithic; BAC, Battle Axe Culture; C, Chalcolithic; TRB, Trichterbecherkultur (Funnelbeaker culture); GAC, Globular Amphora Culture. Original figure and caption from (Ringbauer et al., 2024) figure 4. IBD= Long DNA segments shared between two individuals, known as identity-by-descent (IBD); ancIBD= method for identifying IBD segments in ancient human DNA (aDNA) using a hidden Markov model and imputed genotype probabilities.
However, so enormous is the interpretive advantage gained from the study of ancient DNA that, against all criticism (and there have been and will be many), its very study is now central to any historical archaeological reconstruction that wants to be consistent and useful. Table 1 summarises what I believe to be the advantages and disadvantages of the genetic analysis approach to past populations.

Table 1. Advantages and weaknesses of using ancient DNA analysis.
Furthermore (and here we anticipate the conclusions a little), the presence of ancient DNA analysis is now considered an indispensable step in the study of a past population. Recent papers have shown how combining these studies with isotopic analyses can be extremely advantageous in reconstructing past populations’ histories (Rivollat et al., 2023).
A different approach: the study of individual life histories
The approach to reconstructing individual life histories is a commonly used concept in biology, palaeoanthropology and the social sciences. This approach focuses on the study of an individual's life course in relation to the environment and, in the case of humans, cultural and social contexts (Agarwal, 2016). Unfortunately, beyond the historical sources associated with the emergence of writing, the individual life histories of our ancestors can only be reconstructed indirectly. Reconstructions, even effective ones, of the living conditions of past populations can be obtained from the sensu lato archaeological and bioarchaeological record (i.e. human remains, archaeobotanical and archaeozoological remains) from sediments that have yielded traces of hominid presence/frequency (de Vareilles et al., 2021, Hardy, 2010, Zohar et al., 2022). See Figure 2. However, while it is possible to reconstruct an 'average' life within a group in this way, information on individuals’ lives or their variability is not accessible by these methods. Even in the more recent past, such as in the medieval period, written records are scarce and typically provide information on aspects of elite lives, but provide little to no information on the population as a whole (Fleming, 2009).
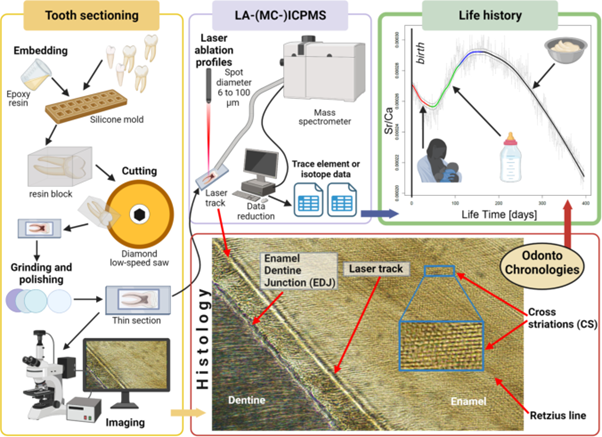
Figure 2. Schematic representation of the analytical workflow for the reconstruction of individual life histories from dental histology coupled with spatially-resolved laser-based biogeochemical analysis. The workflow foresees the tooth sectioning (yellow box). After photographic documentation, the tooth is sectioned using a standard histological protocol (Nava et al., 2019). The thin section is digitally recorded with a high-resolution microscopy digital camera. The photomosaic is analysed to retrieve dental growth parameters. The same thin section is subjected to biogeochemical analysis using a laser ablation instrument coupled to an ICPMS or an MC-ICPMS to measure elemental ratios (Müller et al., 2019) and isotope ratios (Lugli et al., 2022, Müller and Anczkiewicz, 2016) respectively (purple box). The determination of odontochronologies along the laser track allows for the reconstruction of the individual life history during the period of tooth formation, for example, the reconstruction of the early life dietary history (green box). Modified from Esposito et al. (2024)
In contrast, the skeletons of individuals from archaeological or palaeontological contexts provide a repository of information that can be studied using various indicators (morphological, micromorphological and biogeochemical) aimed at extracting, studying, and understanding individual life histories (e.g. Austin et al., 2023, Fleming, 2009, Montgomery, 2010, Nava et al., 2020a, Smith et al., 2021, Müller et al., 2024) and integrating them into the relevant archaeological historical context (Sperduti et al., 2018). Information that can be obtained includes growth and developmental trajectories, physiological stress prevalence during growth, mobility, and diet, including weaning and exposure to toxic substances (see Nava et al., 2024, Müller et al., 2024 for a review). Furthermore, the appositional growth process controlling tooth formation inherently stores a time-stamped signal of such parameters, which—if extracted with a suitably high spatial resolution—also allow for weekly, if not daily, temporal resolution (Nava et al., 2024, Müller et al., 2019). This approach can partially overcome the intrinsic problems of studying mortality samples (as outlined above). Indeed, teeth allow for a close-to-longitudinal analysis, allowing for the study of the individual/environment relationship (health, diet, mobility, pregnancy, puberty) during life and prior to death. See Figure 3. With this approach, we are no longer forced to exclusively analyse the moment of the individual's death, creating models based on (quasi)longitudinal rather than cross-sectional sampling.
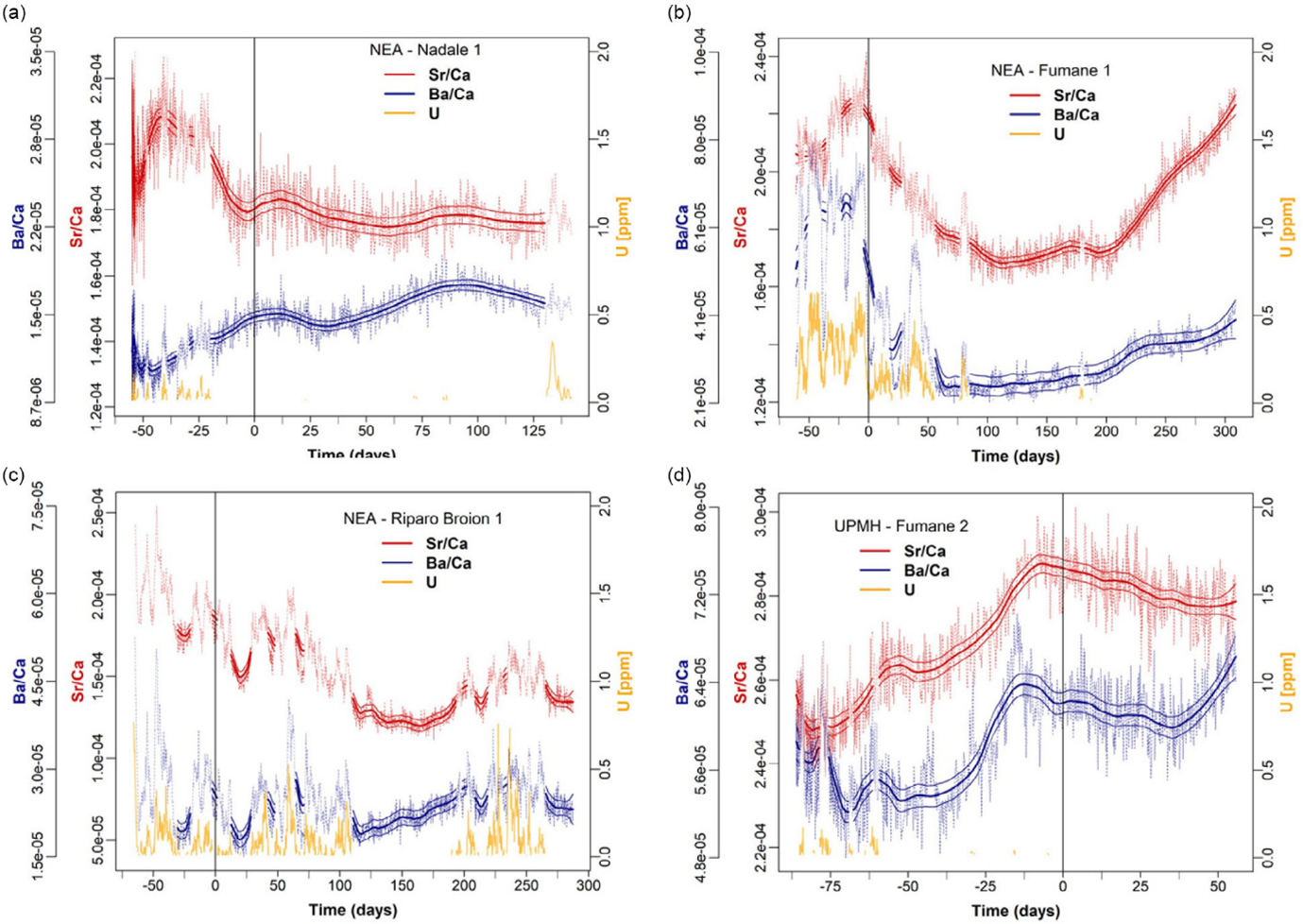
Figure 3. Time-resolved dietary histories based on Sr/Ca Sr variations in Middle-Upper Paleolithic deciduous teeth from Nava et al. (2020b). Elemental profiles (Sr/Ca; Ba/Ca for comparison) were analysed within the enamel closest to the enamel-dentine junction (EDJ); [U] is reported as a diagenetic alteration proxy for all fossil specimens; diagenetically affected sections are grayed out. All are plotted relative to secretion time (in days); the birth event is highlighted by a vertical line in each plot. Elemental ratios are given on a mass (weight) basis, not as mol/mol (Müller et al., 2019). The compositional profiles were smoothed with a locally weighted polynomial regression fit (LOWESS), with the associated SE range (±3 SE) for each predicted value. (a) Nadale 1: the slight decrease in Sr/Ca indicates exclusive breastfeeding until the end of crown formation (4.7 months). (b) Fumane 1: Sr/Ca variation indicates breastfeeding until 4 months of age (fully comparable with modern MCS1 sample not shown here), see Nava et al. (2020). (c) Riparo Broion 1: Sr/Ca profile indicates exclusive breastfeeding until 5 months of age. (d) Fumane 2: 55 days of available postnatal enamel indicates exclusive breastfeeding. The birth event is indicated by a vertical line in each sample. NEA, Neanderthal; UPMH, Upper Palaeolithic Modern Human.
Highly time-stamped reconstructions of individual life histories (Müller et al., 2024) based on high resolution biogeochemical analyses of dental enamel (see Figures 2 and 3) allow a more nuanced and comprehensive understanding of past human behaviour and provide essential information for the understanding of human bio-cultural adaptations. They may also open cognitive windows into aspects such as the mother-infant relationship, pregnancy, lactation, infant stress, the diet of both offspring and mother, high-resolution mobility and many other aspects of the life of past populations that the study of ancient DNA and skeletal morphology cannot provide (Nava, 2024, Nava et al., 2024). To summarize: where ancient DNA is silent, the advanced study of life histories speaks.
The importance of studying infancy and childhood
Over fifty years ago, one of the pillars of post-war Italian biological anthropology (I prefer eluding the name) published a paper in which he stated that the study of the young and infants can tell us so little. To this, I would contend that seldom has greater nonsense been written in the discipline of the bio-cultural study of past human populations. On the contrary, and as has clearly been shown in numerous articles (e.g. Gowland and Halcrow, 2019, Halcrow et al., 2020, Halcrow and Tayles, 2011, Hodgkins et al., 2021, Lugli et al., 2020, Mahoney et al., 2021, McFarlane et al., 2021, Nava et al., 2017, Nava et al., 2019, Nava et al., 2020a, Romandini et al., 2020), the study of infants and juveniles is perhaps one of the most promising research areas in human palaeobiology today.
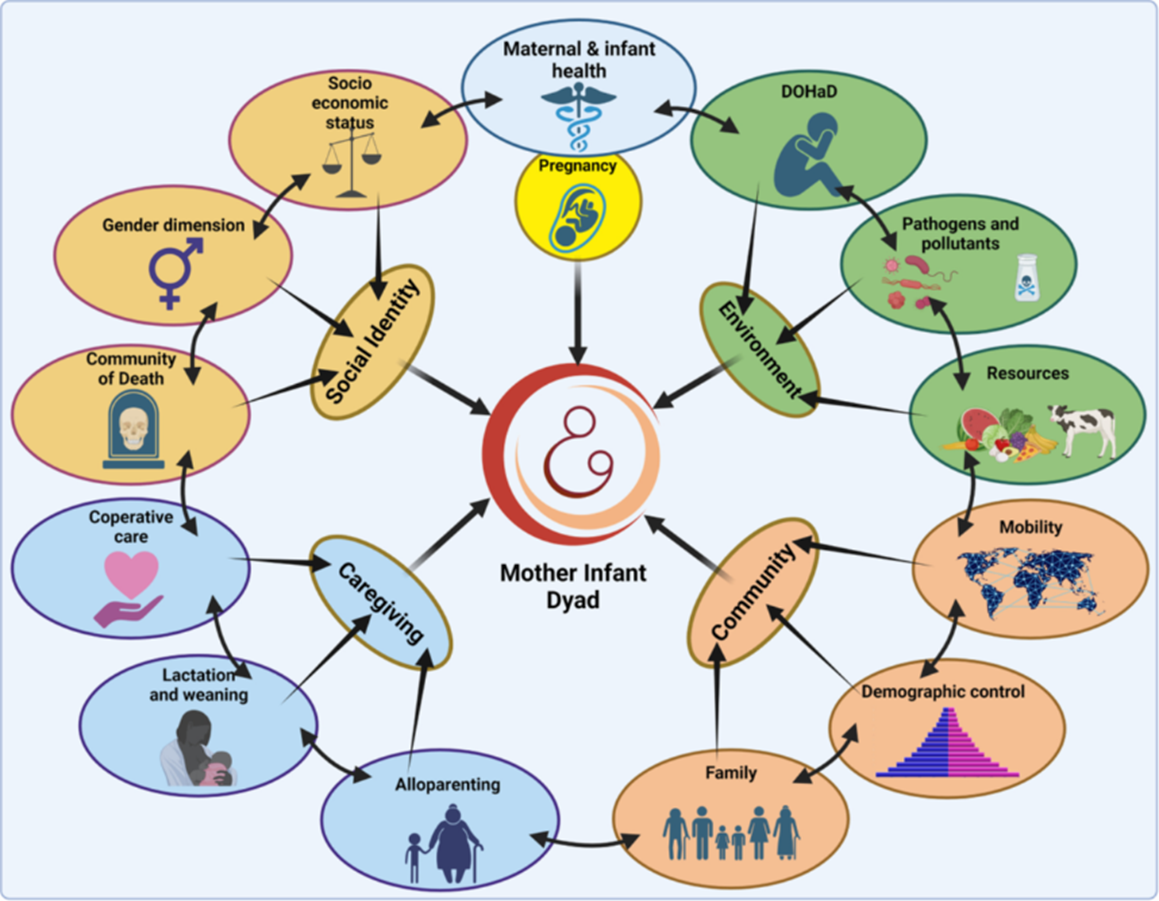
Figure 4. Model of the bio-cultural relationship between mother and children (from Nava et al., 2024).
The choice to study childhood and adolescence/early adulthood is dictated by four things:
- The importance and interest of studying the juvenile portion of an ancient population that represents the true and only projection into the future for a human group. See Figure 4;
- The fact that the study of life history from osteo-dental remains has its greatest informative potential when studying deciduous and permanent teeth (which cover the individual life span from the first weeks of life in utero at ca. 15 gestational weeks to, discontinuously in the latter part, the last 20 years of age; AlQahtani et al., 2010);
- Teeth do not remodel during life (they only deteriorate) and, thus, represent faithful records of individual life (Nava et al., 2019; in this last paper, however, it is shown that a rather unexpected degree of infidelity characterizes the individual dental record. Indeed, the dental crowns of a single individual seems to be not equally susceptible to record physiological stress events, both inter and intra dental crowns);
- Teeth tend to be better preserved than bones and are less affected by diagenesis as dental enamel in its compactness and very low organic component is much less subject to post-depositional chemical-physical changes (Chiaradia et al., 2003, Hoppe et al., 2003, Trickett et al., 2003, Bentley, 2006).
All that glitters is not always gold
The reconstruction of individual life histories, especially for the period of life closest to birth, is a powerful and important tool for accessing the complex world of past human populations. However, it is not a perfect tool. Knowing its problems allows it to be used wisely (if possible) and effectively. In all honesty, it must be admitted that:
- These are always destructive or semi-destructive methods. This is the price we pay (but see Esposito et al. 2024 on how to reduce the damage as much as possible);
- When using non-exfoliated (teeth which have not been lost by the individual through natural replacement with permanent teeth) deciduous teeth, we are always dealing with individuals from funerary or palaeontological contexts. Individuals who died while still young and immature are not representative of the corresponding age segment in the reference population;
- When permanent teeth are used, they must be preserved at their best, which means that heavily worn teeth in which the crown has practically disappeared or greatly reduced are not useful for analysis: it follows that, depending on the research question, only the youngest cohorts will be included in the analysis and the elderly will be substantially excluded. But root cementum analysis is still available for specimens from elderly people: cementum does not have as fine a time resolution as does dentine (~1 year of temporal resolution), but it nevertheless contains a wealth of important information (Cerrito et al., 2020, Cerrito et al., 2022, Nava et al., 2024, Hogg, 2018, Cerrito et al., 2024);
- Among mineralized dental tissues, it is practically only dental enamel which has long-term informative power as it is little affected by diagenesis (Kohn et al., 1999). Dentine and cementum tend to have a biogeochemical and sometimes even histomorphometric signal which could be altered.
The ideal sample: Exfoliated teeth
Within the human palaeobiological record there is a class of remains that can be considered 'ideal' for the study and reconstruction of individual life history: naturally exfoliated deciduous teeth. Such teeth are ideal in that they are intra-vitam lost teeth, i.e. the individual who lost them survived, meaning that such teeth are therefore not affected by the problems associated with mortality specimens. Contrary to what is expected for such a class of remains, they are well represented in the palaeoanthropological record. This is especially true in Italy, where painstaking cave excavations in Middle and Upper Palaeolithic horizons have yielded numerous examples that even represent a majority of the available human dental finds (Alciati et al., 1998, AnzideI et al., 2012, Arnaud et al., 2017, Nava et al., 2020a, Oxilia et al., 2020, Romandini et al., 2020, Villa and Giacobini, 2005, Oxilia et al., 2022).
The virtuous feedback loop: From the population approach to the reconstruction of individual life histories and back again
The reconstruction of life and death patterns in ancient populations from their odontoskeletal remains is a rather delicate, complex and often unachievable task. However, the informative value of bones and teeth as a source of data for evolutionary and historical studies has been increasingly recognized. This type of data is certainly more ‘talkative’ when it is treated with a combined individual/population approach and when it is interpreted within a broader, multidisciplinary framework.
The analytical pipeline that should unite into a single interpretive system both the population and reconstruction of individual life histories approaches has yet to be built in all its complexity, but for some years now there have been examples of noteworthy scientific contributions that approach the study and interpretation of complex burials in an aggregated way. (I do not like the adjective holistic, as I do not believe that a holistic approach sensu stricto will ever be possible). I can only mention a few studies here: for example, the contribution by Tian et al. (2024) represents an excellent study in which archaeology, ancestry, kinship, mortality, and individual mobility are brought together to provide a stronger archaeological-historical interpretive framework. Similarly, the study of the social practices of Avar communities (Gnecchi-Ruscone et al., 2024), the combined use of ancient DNA and radiocarbon dating to increase chronological accuracy (Sedig et al., 2021), the extensive pedigrees analysis to reveal the social organization of a Neolithic community (Rivollat et al., 2023), the use of multiple bioarchaeological methods to face an enigmatic and puzzling Neolithic/Roman Imperial context (Veselka et al., 2024), are all examples of how I think bioarcheological research will develop in the near future. Indeed, the combination of different and transdisciplinary approaches should be the norm in present and future research designs, creating, perhaps, a new revolution in (bio)archaeology.
Conclusions
Technological, and consequently methodological, developments now offer extremely powerful tools that significantly increase the load of information extracted from fossil or archaeological remains.
The hope and aim in writing this text were to bring together a (fairly heterogeneous) set of arguments that all speak one language: that of reconstructing individual life histories in modern bioarchaeological research to be combined with a population approach.
A few months ago, Ian Hodder (Stanford University) gave an interesting lecture at the University of Padua with the intriguing title “From Object and Things to Flows and Waves: an archaeological perspective”. Therein, he proposed (with a complex dialectic) the creation of an approach to the archaeological record that goes beyond the fixity of structuring dates, layers and interactions to a new way of looking at the archaeological record in which flows and waves of change describe the histories of the past.
During the long and articulate discussion at this lecture, I took the liberty of pointing out that this is an approach that is already fully functional in bioarchaeology, where we have moved from the odontoskeletal record described as the fixation in death of certain characteristics of the individual to the reconstruction of dental and skeletal biographies seen as life histories and whose comparison as time (and spatial) series allows us to open hitherto unknown windows into the lives of communities of the past.
It is my hope that this text is a contribution to this fascinating and on-going discussion.
Bibliography
- Agarwal, S. C. 2016. Bone morphologies and histories: Life course approaches in bioarchaeology. American Journal of Physical Anthropology, 159, 130-149.
- Akbari, A., Barton, A. R., Gazal, S., Li, Z., Kariminejad, M., Perry, A., Zeng, Y., Mittnik, A., Patterson, N., Mah, M., Zhou, X., Price, A. L., Lander, E. S., Pinhasi, R., Rohland, N., Mallick, S. & Reich, D. 2024. Pervasive findings of directional selection realize the promise of ancient DNA to elucidate human adaptation. bioRxiv.
- Alciati, G., Coppa, A., Dalmeri, G., Giacobini, G., Lanzinger, M. & Macchiarelli, R. 1998. Human deciduous dental crowns from the Epigravettian layers at Riparo Dalmeri (Trento). A preliminary descriptive note. Preistoria Alpina, 197-200.
- Alqahtani, S. J., Hector, M. & Liversidge, H. 2010. Brief communication: the London atlas of human tooth development and eruption. American Journal of Physical Anthropology, 142, 481-490.
- Anzidei, A. P., Bulgarelli, G. M., Catalano, P., Cerilli, E., Gallotti, R., Lemorini, C., Milli, S., Palombo, M. R., Pantano, W. & Santucci, E. 2012. Ongoing research at the late Middle Pleistocene site of La Polledrara di Cecanibbio (central Italy), with emphasis on human–elephant relationships. Quaternary International, 255, 171-187.
- Arnaud, J., Benazzi, S., Romandini, M., Livraghi, A., Panetta, D., Salvadori, P. A., Volpe, L. & Peresani, M. 2017. A Neanderthal deciduous human molar with incipient carious infection from the Middle Palaeolithic De Nadale cave, Italy. American Journal of Physical Anthropology, 162, 370-376.
- Austin, C., Kumar, P., Carter, E. A., Lee, J., Smith, T. M., Hinde, K., Arora, M. & Lay, P. A. 2023. Stress exposure histories revealed by biochemical changes along accentuated lines in teeth. Chemosphere, 329, 138673.
- Benson, D. A., Cavanaugh, M., Clark, K., Karsch-Mizrachi, I., Lipman, D. J., Ostell, J. & Sayers, E. W. 2013. GenBank. Nucleic Acids Res, 41, D36-42.
- Bentley, R. A. 2006. Strontium isotopes from the earth to the archaeological skeleton: A review. Journal of Archaeological Method and Theory, 13, 135-187.
- Buikstra, J. E. & Ubelaker, D. H. 1994. Standards for data collection from human skeletal remains. Arkansas Archeological Survey Research Series. Fayetteville, AR: Arkansas Archeological Survey.
- Cerrito, P., Bailey, S. E., Hu, B. & Bromage, T. G. 2020. Parturitions, menopause and other physiological stressors are recorded in dental cementum microstructure. Scientific reports, 10, 5381.
- Cerrito, P., Cherian, B. E., Hu, B. & Bromage, T. G. 2024. Population differences in dental cementum growth rates: Implications for using cementum thickness as a method for age estimation. American Journal of Biological Anthropology, 185, e24985.
- Cerrito, P., Nava, A., Radovčić, D., Borić, D., Cerrito, L., Basdeo, T., Ruggiero, G., Frayer, D. W., Kao, A. P., Bondioli, L., Mancini, L. & Bromage, T. G. 2022. Dental cementum virtual histology of Neanderthal teeth from Krapina (Croatia, 130–120 kyr): an informed estimate of age, sex and adult stressors. Journal of the Royal Society Interface, 19, 20210820.
- Chiaradia, M., Gallay, A. & Todt, W. 2003. Different contamination styles of prehistoric human teeth at a Swiss necropolis (Sion, Valais) inferred from lead and strontium isotopes. Applied Geochemistry, 18, 353-370.
- De Vareilles, A., Pelling, R., Woodbridge, J. & Fyfe, R. 2021. Archaeology and agriculture: plants, people, and past land-use. Trends in Ecology & Evolution, 36, 943-954.
- Dewitte, S. N. & Stojanowski, C. M. 2015. The osteological paradox 20 years later: past perspectives, future directions. Journal of Archaeological Research, 23, 397-450.
- Esposito, C., Higgins, O. A., Galbusera, A., Gigante, M., Lugli, F., Müller, W., Madgwick, R., Coppa, A., Benazzi, S., Bondioli, L. & Nava, A. 2024. NOthing goes to WAste (NOWA): A protocol to optimise sampling of ancient teeth. Journal of Archaeological Science, 171, 106087.
- Fleming, R. 2009. Writing Biography at the Edge of History. American Historical Review, 114, 606-614.
- Gnecchi-Ruscone, G. A., Rácz, Z., Samu, L., Szeniczey, T., Faragó, N., Knipper, C., Friedrich, R., Zlámalová, D., Traverso, L. & Liccardo, S. 2024. Network of large pedigrees reveals social practices of Avar communities. Nature, 1-8.
- Gowland, R. & Halcrow, S. 2019. The Mother-infant Nexus in Anthropology: Small Beginnings, Significant Outcomes, Springer.
- Green, R. E., Krause, J., Briggs, A. W., Maricic, T., Stenzel, U., Kircher, M., Patterson, N., Li, H., Zhai, W. & Fritz, M. H.-Y. 2010. A draft sequence of the Neandertal genome. science, 328, 710-722.
- Halcrow, S., Warren, R., Kushnick, G. & Nowell, A. 2020. Care of Infants in the Past: Bridging evolutionary anthropological and bioarchaeological approaches. Evolutionary Human Sciences, 2.
- Halcrow, S. E. & Tayles, N. 2011. The bioarchaeological investigation of children and childhood. Social bioarchaeology, 333-360.
- Hardy, B. L. 2010. Climatic variability and plant food distribution in Pleistocene Europe: Implications for Neanderthal diet and subsistence. Quaternary Science Reviews, 29, 662-679.
- Hodgkins, J., Orr, C. M., Gravel-Miguel, C., Riel-Salvatore, J., Miller, C. E., Bondioli, L., Nava, A., Lugli, F., Talamo, S., Hajdinjak, M., Cristiani, E., Romandini, M., Meyer, D., Drohobytsky, D., Kuester, F., Pothier-Bouchard, G., Buckley, M., Mancini, L., Baruffaldi, F., Silvestrini, S., Arrighi, S., Keller, H. M., Griggs, R. B., Peresani, M., Strait, D. S., Benazzi, S. & Negrino, F. 2021. An infant burial from Arma Veirana in northwestern Italy provides insights into funerary practices and female personhood in early Mesolithic Europe. Scientific Reports, 11, 1-13.
- Hogg, R. 2018. Permanent record: the use of dental and bone microstructure to assess life history evolution and ecology. Methods in paleoecology: Reconstructing Cenozoic terrestrial environments and ecological communities, 75-98.
- Hoppe, K. A., Koch, P. L. & Furutani, T. T. 2003. Assessing the preservation of biogenic strontium in fossil bones and tooth enamel. International Journal of Osteoarchaeology, 13, 20-28.
- Kodama, Y., Shumway, M. & Leinonen, R. 2012. The Sequence Read Archive: explosive growth of sequencing data. Nucleic acids research, 40, D54-D56.
- Kohn, M. J., Schoeninger, M. J. & Barker, W. W. 1999. Altered states: Effects of diagenesis on fossil tooth chemistry. Geochimica et Cosmochimica Acta, 63, 2737-2747.
- Kristiansen, K. 2014. Towards a new paradigm? The third science revolution and its possible consequences in archaeology. Current Swedish Archaeology, 22, 11-34.
- Leinonen, R., Sugawara, H. & Shumway, M. 2011. International nucleotide sequence database C. The sequence read archive. Nucleic Acids Res, 39, D19-D21.
- Lugli, F., Figus, C., Silvestrini, S., Costa, V., Bortolini, E., Conti, S., Peripoli, B., Nava, A., Sperduti, A. & Lamanna, L. 2020. Sex-related morbidity and mortality in non-adult individuals from the Early Medieval site of Valdaro (Italy): the contribution of dental enamel peptide analysis. Journal of Archaeological Science: Reports, 34, 102625.
- Lugli, F., Nava, A., Sorrentino, R., Vazzana, A., Bortolini, E., Oxilia, G., Silvestrini, S., Nannini, N., Bondioli, L. & Fewlass, H. 2022. Tracing the mobility of a Late Epigravettian (~ 13 ka) male infant from Grotte di Pradis (Northeastern Italian Prealps) at high-temporal resolution. Scientific Reports, 12, 1-13.
- Mahoney, P., Mcfarlane, G., Smith, B. H., Miszkiewicz, J. J., Cerrito, P., Liversidge, H., Mancini, L., Dreossi, D., Veneziano, A., Bernardini, F., Cristiani, E., Behie, A., Coppa, A., Bondioli, L., Frayer, D., Radovčić, D. & Nava, A. 2021. Growth of Neanderthal infants from Krapina (120–130 ka), Croatia. Proceedings of the Royal Society B, 288, 20212079.
- Mallick, S., Micco, A., Mah, M., Ringbauer, H., Lazaridis, I., Olalde, I., Patterson, N. & Reich, D. 2024. The Allen Ancient DNA Resource (AADR) a curated compendium of ancient human genomes. Scientific Data, 11, 182.
- Martyn, R., Craig, O. E., Ellingham, S., Islam, M., Fattore, L., Sperduti, A., Bondioli, L. & Thompson, T. 2020. A re-evaluation of manner of death at Roman Herculaneum following the AD 79 eruption of Vesuvius. Antiquity, 94, 76-91.
- Martyn, R., Garnsey, P., Fattore, L., Petrone, P., Sperduti, A., Bondioli, L. & Craig, O. 2018. Capturing Roman dietary variability in the catastrophic death assemblage at Herculaneum. Journal of Archaeological Science: Reports, 19, 1023-1029.
- Mcfarlane, G., Loch, C., Guatelli-Steinberg, D., Bayle, P., Le Luyer, M., Sabel, N., Nava, A., Floyd, B., Skinner, M., White, S., Pitfield, R. & Mahoney, P. 2021. Enamel daily secretion rates of deciduous molars from a global sample of children. Archives of Oral Biology, 132, 105290.
- Montgomery, J. 2010. Passports from the past: Investigating human dispersals using strontium isotope analysis of tooth enamel. Annals of Human Biology, 37, 325-346.
- Müller, W. & Anczkiewicz, R. 2016. Accuracy of laser-ablation (LA)-MC-ICPMS Sr isotope analysis of (bio) apatite–a problem reassessed. Journal of Analytical Atomic Spectrometry, 31, 259-269.
- Müller, W., Lugli, F., Mccormack, J., Evans, D., Anczkiewicz, R., Bondioli, L. & Nava, A. 2024. Human Life Histories. Treatise on Geochemistry 3rd edition.
- Müller, W., Nava, A., Evans, D., Rossi, P. F., Alt, K. W. & Bondioli, L. 2019. Enamel mineralization and compositional time-resolution in human teeth evaluated via histologically-defined LA-ICPMS profiles. Geochimica et Cosmochimica Acta, 255, 105-126.
- Mulligan, C. J. 2021. Systemic racism can get under our skin and into our genes. American Journal of Physical Anthropology, 175, 399-405.
- Nava, A. 2024. Understanding the maternal-infant nexus from dental histology and high-resolution compositional biogeochemistry: implications for bioarchaeological research. Bulletins et Mémoires de la Société d’Anthropologie de Paris, 36.
- Nava, A., Bondioli, L., Coppa, A., Dean, M. C., Rossi, P. F. & Zanolli, C. 2017. New Regression Formula to Estimate the Prenatal Crown Formation Time of Human Deciduous Central Incisors Derived from a Roman Imperial Sample (Velia, Salerno, I-II cent. CE). PloS ONE, 12, e0180104.
- Nava, A., Frayer, D. W. & Bondioli, L. 2019. Longitudinal analysis of the microscopic dental enamel defects of children in the Imperial Roman community of Portus Romae (necropolis of Isola Sacra, 2nd to 4th century CE, Italy). Journal of Archaeological Science: Reports, 23, 406-415.
- Nava, A., Lugli, F., Lemmers, S., Cerrito, P., Mahoney, P., Bondioli, L. & Müller, W. 2024. Reading children’s teeth to reconstruct life history and the evolution of human cooperation and cognition: The role of dental enamel microstructure and chemistry. Neuroscience & Biobehavioral Reviews, 163, 105745.
- Nava, A., Lugli, F., Romandini, M., Badino, F., Evans, D., H, H. A., Gregorio, O., Arrighi, S., Bortolini, E., Delpiano, D., Duches, R., Figus, C., Livraghi, A., Marciani, G., Silvestrini, S., Cipriani, A., Giovanardi, T., Pini, R., Tuniz, C., Bernardini, F., Dori, I., Coppa, A., Cristiani, E., Dean, C., Bondioli, L., Peresani, M., Müller, W. & Benazzi, S. 2020a. Early life of Neanderthals. Proceedings of the National Academy of Sciences of the United States of America, 117 (46) 28719-28726.
- Nava, A., Lugli, F., Romandini, M., Badino, F., Evans, D., Helbling, A. H., Oxilia, G., Arrighi, S., Bortolini, E., Delpiano, D., Duches, R., Figus, C., Livraghi, A., Marciani, G., Silvestrini, S., Cipriani, A., Giovanardi, T., Pini, R., Tuniz, C., Bernardini, F., Dori, I., Coppa, A., Cristiani, E., Dean, C., Bondioli, L., Peresani, M., Muller, W. & Benazzi, S. 2020b. Early life of Neanderthals. Proc Natl Acad Sci U S A, 117, 28719-28726.
- Orlando, L., Allaby, R., Skoglund, P., Der Sarkissian, C., Stockhammer, P. W., Ávila-Arcos, M. C., Fu, Q., Krause, J., Willerslev, E. & Stone, A. C. 2021. Ancient DNA analysis. Nature Reviews Methods Primers, 1, 14.
- Oxilia, G., Bortolini, E., Badino, F., Bernardini, F., Gazzoni, V., Lugli, F., Romandini, M., Radini, A., Terlato, G. & Marciani, G. 2020. Exploring late Paleolithic and Mesolithic diet in the Eastern Alpine region of Italy through multiple proxies. American Journal of Physical Anthropology.
- Oxilia, G., Bortolini, E., Marciani, G., Menghi Sartorio, J. C., Vazzana, A., Bettuzzi, M., Panetta, D., Arrighi, S., Badino, F., Figus, C., Lugli, F., Romandini, M., Silvestrini, S., Sorrentino, R., Moroni, A., Donadio, C., Morigi, M. P., Slon, V., Piperno, M., Talamo, S., Collina, C. & Benazzi, S. 2022. Direct evidence that late Neanderthal occupation precedes a technological shift in southwestern Italy. American Journal of Biological Anthropology, 179, 18-30.
- Peter, B. M., Petkova, D. & Novembre, J. 2020. Genetic landscapes reveal how human genetic diversity aligns with geography. Molecular biology and evolution, 37, 943-951.
- Prüfer, K., De Filippo, C., Grote, S., Mafessoni, F., Korlević, P., Hajdinjak, M., Vernot, B., Skov, L., Hsieh, P. & Peyrégne, S. 2017. A high-coverage Neandertal genome from Vindija Cave in Croatia. Science, 358, 655-658.
- Prüfer, K., Racimo, F., Patterson, N., Jay, F., Sankararaman, S., Sawyer, S., Heinze, A., Renaud, G., Sudmant, P. H. & De Filippo, C. 2014. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature, 505, 43-49.
- Reich, D. 2018. Who we are and how we got here: Ancient DNA and the new science of the human past, Oxford University Press.
- Ringbauer, H., Huang, Y., Akbari, A., Mallick, S., Olalde, I., Patterson, N. & Reich, D. 2024. Accurate detection of identity-by-descent segments in human ancient DNA. Nature Genetics, 56, 143-151.
- Rivollat, M., Rohrlach, A. B., Ringbauer, H., Childebayeva, A., Mendisco, F., Barquera, R., Szolek, A., Le Roy, M., Colleran, H. & Tuke, J. 2023. Extensive pedigrees reveal the social organization of a Neolithic community. Nature, 620, 600-606.
- Romandini, M., Oxilia, G., Bortolini, E., Peyrégne, S., Delpiano, D., Nava, A., Panetta, D., Di Domenico, G., Martini, P., Arrighi, S., Badino, F., Figus, C., Lugli, F., Marciani, G., Silvestrini, S., Menghi Sartorio, J. C., Terlato, G., Hublin, J.-J., Meyer, M., Bondioli, L., Higham, T., Slon, V., Peresani, M. & Benazzi, S. 2020. A late Neanderthal tooth from northeastern Italy. Journal of Human Evolution, 147, 102867.
- Schroeder, L. 2020. Revolutionary fossils, ancient biomolecules, and reflections in ethics and decolonization: paleoanthropology in 2019. American Anthropologist, 122, 306-320.
- Sedig, J. W., Olalde, I., Patterson, N., Harney, E. & Reich, D. 2021. Combining ancient DNA and radiocarbon dating data to increase chronological accuracy. Journal of Archaeological Science, 133, 105452.
- Smith, T. M., Cook, L., Dirks, W., Green, D. R. & Austin, C. 2021. Teeth reveal juvenile diet, health and neurotoxicant exposure retrospectively: What biological rhythms and chemical records tell us. BioEssays, 2000298.
- Soncin, S., Talbot, H. M., Fernandes, R., Harris, A., Von Tersch, M., Robson, H. K., Bakker, J. K., Richter, K. K., Alexander, M., Ellis, S., Thompson, G., Amoretti, V., Osanna, M., Caso, M., Sirano, F., Fattore, L., Colonese, A. C., Garnsey, P., Bondioli, L. & Craig, O. E. 2021. High-resolution dietary reconstruction of victims of the 79 CE Vesuvius eruption at Herculaneum by compound-specific isotope analysis. Science Advances, 7, eabg5791.
- Sperduti, A., Bondioli, L., Craig, O. E., Prowse, T., Garnsey, P., & Scheidel, W. 2018. Bones, teeth and history. In W. Scheidel ed. The Science of Roman History: Biology, climate and the future of the past. Princeton Univ. Press 123-173.
- Sperduti, A., D'Agostino, B., Gastaldi, P., Faiella, I., Fiore, I., Pellegrino, C., Rizzo, C. & Bondioli, L. 2021. When children mark the change: funerary rituals and socio-demographic dynamics in Pontecagnano (Salerno, Campania) between the 9th and 5th centuries BCE. Childhood in the Past, 14, 125-144.
- Trickett, M. A., Budd, P., Montgomery, J. & Evans, J. 2003. An assessment of solubility profiling as a decontamination procedure for the Sr-87/Sr-86 analysis of archaeological human skeletal tissue. Applied Geochemistry, 18, 653-658.
- Veselka, B., Reich, D., Capuzzo, G., Olalde, I., Callan, K., Zalzala, F., Altena, E., Goffette, Q., Ringbauer, H. & Van Der Velde, H. 2024. Assembling ancestors: the manipulation of Neolithic and Gallo-Roman skeletal remains at Pommerœul, Belgium. Antiquity, 98, 1576-1591.
- Villa, G. & Giacobini, G. 2005. Nuovi denti umani dai livelli epigravettiani di Riparo Dalmeri (TN). Preistoria Alpina, 41, 245-250.
- Weaver, D. S., Perry, G. H., Macchiarelli, R. & Bondioli, L. 2000. A surgical amputation in 2nd century Rome. The Lancet, 356, 686.
- Wood, J. W., Milner, G. R., Harpending, H. C., Weiss, K. M., Cohen, M. N., Eisenberg, L. E., Hutchinson, D. L., Jankauskas, R., Cesnys, G. & Česnys, G. 1992. The osteological paradox: problems of inferring prehistoric health from skeletal samples [and comments and reply]. Current Anthropology, 33, 343-370.
- Yuan, D., Ahamed, A., Burgin, J., Cummins, C., Devraj, R., Gueye, K., Gupta, D., Gupta, V., Haseeb, M. & Ihsan, M. 2024. The European nucleotide archive in 2023. Nucleic Acids Research, 52, D92-D97.
- Zohar, I., Alperson-Afil, N., Goren-Inbar, N., Prévost, M., Tütken, T., Sisma-Ventura, G., Hershkovitz, I. & Najorka, J. 2022. Evidence for the cooking of fish 780,000 years ago at Gesher Benot Ya’aqov, Israel. Nature Ecology & Evolution, 6, 2016-2028.
Go back to top